HETEROSIS HETEROSIS
(from the Greek heteroiosis - change, transformation), “hybrid power”, the superiority of hybrids in a number of characteristics and properties over the parent forms. The term "G." proposed by J. Schell in 1914. As a rule, G. is characteristic of first-generation hybrids obtained by crossing unrelated forms: decomp. lines, breeds (varieties) and even species. In further generations (crossing hybrids with each other), its effect weakens and disappears. The hypothesis of “overdominance”, or monogenic G., assumes that heterozygotes by definition. gene are superior in their characteristics to the corresponding homozygotes. A phenomenon that illustrates this hypothesis is interallelic complementation. A number of other hypotheses are based on the assumption that the hybrid has a larger number of dominant alleles of different genes compared to the parental forms and the interaction between these alleles. Synthetic hypotheses are based on both intragenic and intergenic interactions. The importance of heterozygosity as the basis of genetics is also evidenced by the fact that in natural populations individuals are heterozygous for a large number of genes. Moreover, many remain in the heterozygous state. alleles that, when homozygous, exhibit adverse effects on vital signs. G. is important in agriculture. practice (in agricultural animals and plants, corn often leads to an increase in productivity and yield: the production of simple and double interline hybrids of corn made it possible to increase the gross grain yield by 20-30%), but its use is often not effective enough, because The problem of consolidating G. in a number of generations has not yet been solved. Vegetative propagation of heterotic forms, polyploidy, etc. are considered as approaches to solving this problem. irregular forms of sexual reproduction (apomixis, parthenogenesis, etc.).
.(Source: “Biological Encyclopedic Dictionary.” Editor-in-chief M. S. Gilyarov; Editorial Board: A. A. Babaev, G. G. Vinberg, G. A. Zavarzin and others - 2nd ed., corrected - M.: Sov. Encyclopedia, 1986.)
heterosis(hybrid power, hybrid strength), superiority of first generation hybrids over parental forms in viability, yield, fertility and a number of other characteristics. To obtain the effect of hybrid power, it is important to choose unrelated forms as parents, representing different lines, breeds, even species. In practice, the best parental pairs, which produce the most valuable hybrids, are selected as a result of numerous crossings, which make it possible to identify the most successful compatibility of different lines. When successive generations are crossed with each other, heterosis weakens and dies out.
Heterosis is based on a sharp increase in heterozygosity in first generation hybrids and superiority heterozygotes for certain genes over the corresponding ones homozygous. Thus, the phenomenon of hybrid power is opposite to the result of inbreeding - inbreeding which has adverse consequences for the offspring. The genetic mechanism of heterosis (it is not fully understood) is also associated with the presence in the hybrid, compared to the parents, of a larger number of dominant genes that interact with each other in a favorable direction.
Heterosis is widely used in agricultural practice to increase agricultural productivity. crops and agricultural productivity animals. In the 1930s US breeders have dramatically increased corn yields by using hybrid seeds. One of the important tasks selection– search for ways to “consolidate” heterosis, i.e. preserving it over generations.
.(Source: “Biology. Modern illustrated encyclopedia.” Chief editor A. P. Gorkin; M.: Rosman, 2006.)
Synonyms:
See what "HETEROSIS" is in other dictionaries:
Acceleration of growth, increase in size, increase in vitality and fertility of first generation hybrids compared to the parental forms of plants or animals. Typically, heterosis subsides in the second and subsequent generations. Heterosis is widespread... ... Financial Dictionary
Heterosis (translated from Greek as change, transformation) is an increase in the viability of hybrids due to the inheritance of a certain set of alleles of various genes from their dissimilar parents. This phenomenon is the opposite of inbred... Wikipedia
- (from the Greek heteroiosis change, transformation), the property of first-generation hybrids to surpass the best of the parent forms in vitality, fertility and other characteristics. In the second and subsequent generations, heterosis usually fades out.... ... Modern encyclopedia
In the breeding of animals and plants, a special place is occupied by the phenomenon of hybrid power, or heterosis, which is as follows.
When crossing different races, animal breeds and plant varieties, as well as inbred lines, F 1 hybrids are often superior to the original parent organisms in a number of traits and properties. Crossing hybrids with each other leads to the attenuation of this effect in subsequent generations. Currently, heterosis has been established for all studied species.
Although the effect of heterosis has been known since ancient times, its nature still remains poorly understood. The first interpretation of the biological significance of heterosis in the evolution of animals and plants and an attempt to explain the mechanism of this phenomenon belonged to Charles Darwin. He systematized a large number of facts and himself conducted experiments to study heterosis. According to Charles Darwin, heterosis is one of the reasons for the biological usefulness of crossing in the evolution of species. Cross-fertilization is supported by natural selection precisely because it serves as a mechanism for maintaining the greatest heterosis,
A deep scientific analysis of the phenomenon of heterosis became possible only from the beginning of the 20th century. after the discovery of basic genetic patterns. When considering heterosis, we will touch upon its occurrence when crossing predominantly inbred lines of the same species, since in this case it is easier to elucidate its genetic mechanism.
Since the beginning of this century, systematic studies of crosses between inbred lines have been carried out on maize. At the same time, G. Schell showed that crossing some lines produces hybrid plants that are more productive in terms of grain and vegetative mass than the original lines and varieties. Experimental data are presented showing the low yield of inbred lines, a significant increase in yield in F 1 and a decrease in F 2 during self-pollination of F 1 plants.
Over the past 30 years, in a number of countries, including ours, sowing with hybrid seeds has become the main method of producing corn both for grain and for silage. To obtain hybrid seeds, first inbred lines are created from the best varieties that meet the requirements of a given climatic region (an inbred line is created within 5-6 years by self-pollination). When selecting lines, their qualities and properties that need to be obtained from the future hybrid organism are assessed. Inbreeding within lines cannot be effective unless it is accompanied by selection.
Having created a large number of inbred lines, they begin crossing between them. Interline hybrids of the first generation are assessed by heterosis effect; Based on this indicator, lines with the best combinational value are selected and then propagated on a large scale to produce hybrid seeds. Work on creating inbred lines and assessing their combinational value is constantly carried out in breeding institutions. The more valuable lines are created, the sooner the best hybrid combinations with the required combination of properties can be found.
When obtaining hybrid seeds for production purposes, the original lines that give the greatest heterosis effect when crossed are sown in rows, alternating maternal and paternal forms. To ensure pollination between them, male inflorescences (panicles) are removed from the mother plants. Now a new scheme for the production of hybrid seeds using cytoplasmic male sterility has been developed, which has significantly reduced the labor required to remove panicles from maternal plants. In this way, simple interline corn hybrids are obtained. This method is, in principle, general for seed production of hybrids of various cross-pollinating plants.
At present, simple interline corn hybrids are not used in agricultural practice, since the costs of obtaining such seeds are not recouped. Sowing seeds of double interline hybrids is now being widely introduced into practice. The latter are obtained by crossing two simple hybrids exhibiting heterosis.
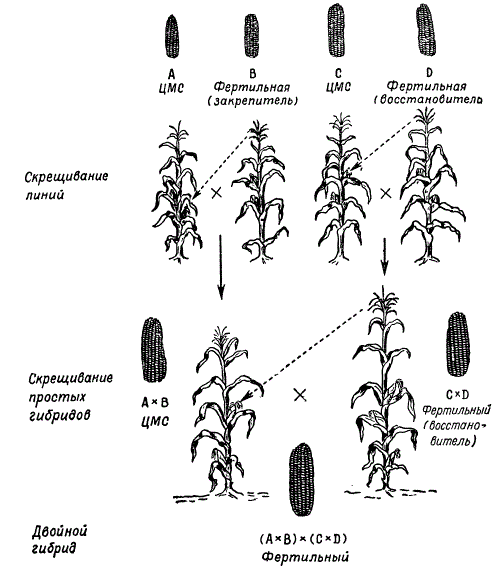
According to M.I. Khadzhinov and G.S. Galeev, the results of assessing the productivity of different hybrids - intervarietal, varietal and double interlinear - show that double interlinear hybrids are the most productive.

The selection of simple hybrids to obtain the most productive double hybrids is an important stage in their selection. The best results are obtained by crossing lines originating from different varieties. So, for example, if one simple hybrid is obtained from crossing inbred lines of two varieties A X B, and another from crossing lines of other varieties C x D, then a double hybrid (A X B) x (C X D) gives heterosis more often than if a double hybrid would be obtained from crossing simple hybrids descended from lines of the same variety: (A x A 1 x (A 2 x A 3) or (B x B 1) x (B 2 x B 3).
For the successful development of hybrid seed production, it is necessary to study, firstly, how long inbreeding should be carried out in order to obtain homozygous lines, and, secondly, to develop methods for more quickly assessing their combinational value.
All that has been said in relation to corn seems to be general for the production of hybrids in other higher cross-pollinating plants, as well as in animals. Currently, in poultry and pig farming in a number of countries, the crossing of inbred lines originating from the same or different breeds is widely used. It should be especially emphasized that the widespread use of hybrids in livestock breeding is possible only with a high level of breeding work and the presence of valuable breeds, it is clear that in most cases, inbred lines, on average, will always have lower performance than varieties. The presence of heterosis should be discussed only in the case when the interline hybrid is superior not only to the parents (lines), but also to the varieties or breeds from which these lines originated.
As we know, genes determine the properties of an organism at all stages of ontogenesis, starting from the moment of fertilization. The genes of the oocyte nucleus are capable of determining the properties of the cytoplasm of the egg even before fertilization. The nature of the implementation of the genotype changes depending on the properties of the cytoplasm of the zygote. Thus, the manifestation of heterosis in a hybrid also depends on the properties of the cytoplasm. The role of the cytoplasm in determining heterosis is proven as follows. When reciprocally crossing two lines A x B and B x A, heterosis for the same properties often appears in hybrids of only one of the crosses and does not appear in hybrids of the other.
The manifestation of heterosis is due to the individual development of the hybrid. During ontogenesis it is realized unevenly. At some stages of ontogenesis, heterosis manifests itself according to some characteristics, and at others - according to others or only according to some. Thus, at an early age, the same hybrid may exhibit heterosis in relation to the growth rate of individual parts of the body and increased resistance to diseases, but it may not exist, for example, in relation to resistance to unfavorable temperatures. Heterosis due to this property may appear later.
The manifestation of heterosis is also strongly influenced by environmental factors in which the hybrid organism develops. There is reason to believe that heterosis in terms of viability and resistance to diseases is better detected under environmental conditions unfavorable for development. Even Charles Darwin suggested that heterosis in hybrids is due to a wider norm of adaptive reactions. Research in recent years has shown that F 1 hybrids, in comparison with inbred lines, are characterized by a greater variety of metabolic processes, the participation of a larger number of different metabolites, growth substances and enzymes. Heterosis manifests itself not only in the whole organism, but also at the cellular level.
Elucidation of the genetic mechanisms of heterosis remains an unresolved problem. Currently, there are three hypotheses trying to explain the occurrence of heterosis:
- heterozygous state for many genes,
- interaction of dominant favorable genes,
- overdominance - a heterozygote is superior to homozygotes.
As we have already said, when crossing homozygous inbred lines, first-generation hybrids acquire a heterozygous state for many genes. In this case, the effect of harmful recessive mutant alleles is suppressed by the dominant alleles of both parents. Schematically, this can be represented as follows: one inbred line in a homozygous state has a recessive allele of one gene aaBB, and the second - another gene AAbb. Each of these recessive gene alleles in the homozygous state determines some deficiency that reduces the viability of the inbred line. When crossing lines aaBB X AAbb, the hybrid combines the dominant alleles of both genes (AaBb). F 1 hybrids will show not only heterosis, but also uniformity in the indicated genes. In F 2, the number of individuals with two dominant genes in the heterozygous state will be only 4/16, so not all individuals are heterotic. In further generations, the number of heterozygotes decreases, and the number of homozygotes increases. For these reasons, heterosis fades in subsequent generations. This is the scheme of one of the genetic hypotheses of heterosis.
We gave an example with two genes, but physiological properties are determined by a huge number of genes. In addition, it is incorrect to assume that dominant alleles always have a positive effect, and recessive ones have a negative effect on the viability of the organism and its productivity. Dominant wild-type alleles are more likely to have a beneficial effect than recessive ones. This can be explained by the fact that unfavorable dominant mutations are subject to more stringent selection - they are already eliminated in the zygote and at the embryonic stages, and only favorable ones are retained by selection. And since the very property of gene dominance evolves under the control of selection, wild-type alleles turn out to be more favorable for the organism. Therefore, the selection of dominant alleles in a hybrid combination can more likely ensure heterosis. If heterosis were caused by a simple set of dominant alleles present in the population, then this set would be easy to compose through a series of crosses and obtain heterotic combinations. It is very possible that some breeds and varieties were developed precisely by crossing and combining lines with a set of favorable dominant alleles. But so far it has not been possible to fix heterosis in the hybrid combination of F 1, that is, to obtain forms that do not split into F 2.
D. Jones proposed an addition to the stated hypothesis back in 1917. According to D. Jones, different genes that give heterosis in combination are located within the same group of linkages: for example, AbcdE - in one, and, accordingly, aBCDe - in another homologous chromosome. It is this circumstance, from the point of view of D. Jones, that makes it difficult to select completely homozygous forms in F 2 for favorable dominant genes that give the combination heterosis. To implement such a combination of dominant alleles, it is necessary that in this chromosome pair AbcdE//aBCDe a minimum of double crossing over occurs, which would lead to the emergence of a chromosome with only dominant ABCDE alleles. The low probability of simultaneous occurrence of such an event in several chromosomes is, according to D. Jones, the reason that complicates the consolidation of heterosis. The F 1 hybrid contains all these dominant alleles, and therefore it exhibits heterosis.
This is the scheme for explaining heterosis according to the second hypothesis - the hypothesis of the interaction of dominant favorable genes, which is sometimes also called the hypothesis of a set of cumulative dominant alleles, and heterosis itself is called mutational. This hypothesis is generally based on the idea of a simple summation of the effect of dominant alleles with a complementary effect.
The third hypothesis is based on the fact that the heterozygous state of alleles is superior to the homozygous one (AA< Aa >aa). Here we can assume a favorable transposition of wild-type and mutant alleles, which somehow enhances the effect of the genes. This explanation of heterosis is called the overdominance hypothesis.
None of the three hypotheses can be considered the only correct one. Now it is premature to give preference to one or the other. They will probably all turn out to be correct, but for different cases. It is likely that each of the mechanisms envisioned by these hypotheses plays a role in determining hybrid power. Heterosis is a complex phenomenon both in terms of the mechanism of its occurrence and its manifestation in ontogenesis.
Obviously, the final conclusion about the genetic mechanisms of heterosis can be made only after the picture of the interaction of genes in the genetic system at the biochemical and molecular levels is revealed. As we already know, suppressing the manifestation of mutations with the help of suppressors allows us to correct disturbances in the action of the gene and its alleles. It is possible that the phenomenon of heterosis is the total result of the work not of the main genes that determine the development of the characteristics of the organism, but of a set of suppressive genes in the genotype. An important point in the study of heterosis is the study of plasma relations.
The main task of using heterosis in breeding is to consolidate it, that is, to preserve the effect of heterosis during the reproduction of the hybrid. The solution to this problem is conceived in several aspects: firstly, the consolidation of heterosis by transferring the hybrid organism from sexual reproduction to apomictic, which is apparently possible for some plants; secondly, the consolidation of heterosis by transferring a diploid hybrid exhibiting heterosis to a polyploid state. In this case, the heterozygous combination of genes will persist longer.
In vegetatively propagated plants, the maintenance of valuable hybrid combinations obtained sexually is carried out by vegetative propagation (cuttings, grafting, tubers, etc.).
There are a number of other ways to maintain heterosis over a number of generations, but all of them have not yet been sufficiently developed and tested.
The method is intended for use in agriculture. The goal is to significantly increase the efficiency of crop hybridization. Heterosis of hybrids appears only in the first generation. It has been established that the attenuation of heterosis in subsequent generations of the hybrid is mainly due to the transition of recessive lethals, semilethals and subvitals into a homozygous state and disruption of a complex of favorable coordinately acting genes. Elimination of these phenomena leads to the consolidation of heterosis in subsequent generations. It is carried out by backcrossing a hybrid with artificially obtained absolutely homozygous androgenetic sons, after which the genetically transformed hybrid is almost completely cleared of harmful genes and at the same time preserves intact the complex of favorable genes that determine heterosis. This makes it possible to completely preserve heterosis in subsequent industrial generations obtained as a result of simple intrahybrid crossings, which has been proven by experiments on silkworms. The method is also intended for agricultural plants, in which it is possible to obtain androgenetic absolutely homozygous individuals. 3 salary, 4 ill.
The invention relates to methods used in agriculture. A well-known natural way of preserving heterosis in subsequent generations is through vegetative propagation in plants that additionally have sexual reproduction. Numerous studies in this area on other plants that are not capable of vegetative propagation and animals were not completely successful (1), because the nature of heterosis was still a big genetic mystery (2). The literature has not even expressed any real theoretical approaches to a radical solution to this important problem. In some animals, heterosis can be fixed by cloning. However, only a few offspring identical to the mother are obtained using this method. In the silkworm, cloning has been developed more successfully, but for practical use in terms of preserving heterosis, it is not acceptable for two reasons: due to the high labor intensity of mass production of parthenogenetic offspring and the lower productivity of the female sex, which makes up the clones, compared to males (3 ). Promising results were obtained by the authors after they developed the method of meiotic parthenogenesis and obtained absolutely homozygous male silkworms from parthenogenetic clones (4). Their backcrossing with a parthenogenetic clone of hybrid origin made it possible to consolidate heterosis in backcross generations (5). But this was the discovery of only the fundamental possibility of fixing heterosis. This method had no practical significance and, therefore, could not be patented as a method. This was explained by the fact that homozygous males could only be obtained from highly viable female parthenoclones with a high propensity for parthenogenesis. It was practically impossible to obtain absolute homozygotes in commercial breeds and hybrids, so meiotic parthenogenesis was used only for reconnaissance experiments aimed at determining the possibility of solving the problem. The invention of a method for fixing heterosis in the silkworm, suitable for production, became possible after the authors discovered monospermic androgenesis (1998, unpublished). The essence of the invention. Heterosis appears only in the first generation of the hybrid. In subsequent generations, starting from the second, it sharply fades. Therefore, in order to grow a heterotic hybrid, it is necessary to repeat intervarietal or interbreed hybridization each time. This process is technically complex and very labor-intensive, and for many plant crops it is simply not feasible, although their hybrids, if they were obtained, would give surprisingly high yields compared to the parent forms. This is exemplified by many agricultural plants. These problems would be radically solved if it were possible to develop an effective way to consolidate heterosis in subsequent generations. This method would simultaneously open up a completely new approach to creating hybrids that are even more outstanding in heterosis. It is known that any industrial hybrid is obtained by crossing a huge mass of individuals of two parental forms. And these individuals are very differentiated in their combinative ability. Therefore, production is content with average heterosis for all individual hybrids taken together, each of which comes from the germ cells of two parents. While rare individual hybrids have truly fantastic heterosis, it is irretrievably lost in the next generation. The proposed method will make it possible to consolidate this powerful heterosis in subsequent generations of the hybrid and propagate it in unlimited quantities. One of the reasons for heterosis was considered to be the beneficial effect on the development and vital activity of the organism of heterozygosity of all genes in general, regardless of their specificity (the “overdominance” hypothesis). Using silkworms, the authors experimentally proved that heterosis occurs as a result of two main reasons. The first is the integration in the genotype of hybrids of a large number of favorable genes, coordinated in their action, that control viability. The second is the transition to a heterozygous state not of all genes of the genotype, but only of recessive details, hemilethals and subvitals (4). In fig. 1 provides evidence of this. Consequently, the decrease in heterosis in subsequent generations of hybrids is mainly explained by the inevitable transition of some recessive parts and semi-lethals into a homozygous state when crossing a hybrid within its limits and the disruption of a complex of favorable genes that increase viability during meiosis. Therefore, the authors came to the conclusion that it is possible to consolidate heterosis in subsequent generations if the complex of all favorable genes in the hybrid genotype is completely preserved or even improved and the recessive lethal and semi-lethal ones are almost completely removed from the genotype. This problem was solved by the authors in the following way. Two genetically distant breeds are selected as the starting material, from crossing which the most highly heterotic hybrids arise. These two breeds produce a series of individual hybrids, each of which comes from only two parents. Through comparative tests, the 10 best individual hybrids in terms of heterosis are selected. From each hybrid, absolutely homozygous descendants are obtained by the method of monospermic androgenesis, which is available to breeders. For this purpose, uninseminated females of any breed are irradiated with rays at a dose of 80 kr. The females then mate with males of the individual hybrids. Layed eggs at the age of 60-80 minutes after laying at a temperature of 25 o C are heated for 210 minutes in water heated to 38 o C. The overwhelming majority of absolute homozygotes die at different stages of development due to the fact that in the haploid genotype they inherited from father, contains many lethal, semi-lethal and subvital genes. When the pronucleus nucleus diploidizes, they pass into a homozygous state, which is most often incompatible with the normal development of the organism. Only those homozygotes survive who, during meiosis, did not receive or received, but very few, harmful genes, mostly of weak effect (5). The grown absolutely homozygous individuals are backcrossed with the original hybrid, thus obtaining the first backcross generation (Fig. 2). The maturation of the original hybrid and absolute homozygotes must be synchronized by delaying the start of growing the first for a time equal to the duration of the development cycle of the selected object. Simple calculations show that in backcross offspring new homozygotes of genes with strong harmfulness cannot appear, and homozygotes of subvital genes, if they were not eliminated from the surviving homozygous androgens, are suppressed by a complex of favorable genes inherited from the original hybrid. This is why heterosis persists in all backcross generations (Fig. 3). The first and subsequent backcross generations are dealt with in exactly the same way as with the original hybrid (Fig. 2). Further backcrosses lead, firstly, to the almost complete removal of details and semi-lethals from the hybrid genotype and, secondly, to the preservation of the numerically predominant part of the genes that provided heterosis in the original hybrid. After 5 or 6 backcrosses, the hybrid, cleared of harmful genes, is massively propagated by intrahybrid crossing. In the offspring obtained as a result of such reproduction, heterosis not only remains at the level of the original hybrid, but even increases somewhat (Fig. 4), which indicates a complete solution to the problem of fixing heterosis in the silkworm. The complete commonality of the genetic basis of heterosis and its attenuation in animals and plants allows this invention to be recommended for consolidating heterosis in agricultural plants, in which it is possible to obtain absolutely homozygous individuals of androgenic origin from hybrids. They are obtained by stimulating the embryonic development of haploid pollen, followed by the transformation of its germ cells into diploid ones, which develop into viable fertile plants. The technique varies depending on the biological characteristics of the crop. Graphic materials. Fig. 1 A. A direct relationship is shown between the yield of silkworm cocoons - the main indicator of heterosis (1) and the levels of heterozygosity (2) of genetic variants of the hybrid that are not purified from lethals and semi-lethals. The yield and heterozygosity indicators of the original hybrid of the first option (1) are taken as 100%. B. It has been shown that there is a complete absence of dependence between the cocoon yield (1) and the levels of heterozygosity (2) in genetic variants purified from lethals and semi-lethals. This proves the inconsistency of the “overdominance” hypothesis of heterosis and the possibility of maintaining heterosis in backcross generations. Fig. 2. Scheme for purifying silkworm hybrids from recessive lethals and semi-lethals through backcrossing of hybrids with absolutely homozygous males of breed A and B obtained from them. F 1, F 2 - hybrid of the first and second generation. F b1, F b2 - first and second backcross generations. Fig. 3. Viability of the original hybrid (1) and backcross generations (II), obtained according to the scheme presented in Fig. 2. Fig. 4. Demonstrates indicators of the frequency of harmful genes in the heterozygous state (1), cocoon mass (2), viability (3) in the original hybrid (I) and the transformed hybrid after four consecutive backcrosses with homozygous males (II), as well as in three consecutive inbred generations (III-V). Each genetic variant was reared simultaneously with a control parthenogenetic hybrid, the indicators of which were taken as 100%. In all genetic variants, heterosis is higher than in the original hybrid, which indicates a radical solution to the problem of fixing heterosis. The stable preservation of heterosis in all backcross generations has already indicated the fundamental effectiveness of the developed method. But backcross generations are not practical due to the difficulty of obtaining them. Therefore, in the final experiment on silkworms, they studied the possibility of fixing heterosis not in backcross, but in normal generations. In this final experiment, the original hybrid was first subjected to four backcrosses with homozygous males. As a result, the frequency of heterozygotes for lethals and semi-lethals decreased to 6.2% from 100% in the original material. Next, backcross generations were propagated by inbreeding. Each inbred generation was obtained by crossing a brother with a sister within each individual family. As a result, the frequency of harmful genes suppressed by normal alleles decreased in the first inbred generation to 4.7, and in the second and third - to 3.5 and 2.6%, respectively. Inbred reproduction has an extremely detrimental effect on all economic indicators of normal inbred offspring. But in our experiment, it not only did not have a depressing effect on the inbred offspring, but, on the contrary, led to an increase in its average weight of one cocoon and viability compared to the original, control hybrid (Fig. 4). Consequently, the problem of fixing heterosis in hybrids of subsequent generations has been radically solved. BIBLIOGRAPHICAL DATA
1. Inge-Vechtomov S.I. 1989. Genetics with the basics of selection. M. "Higher School", on page 557. 2. Hutt F. 1969. Animal genetics. Per. from English edited by Doctor of Biology Sciences Ya.L. Glembotsky. M., "Spike", on page 322. 3. Strunnikov V. A. 1998. Cloning of animals: theory and practice. - Nature, N 7, pp. 3 -9. 4. Strunnikov V.A. 1987. Genetic methods for selection and sex regulation of silkworms. M. VO "Agropromizdat", on page 35. 5. Strunnikov V.A. 1994. The nature of heterosis and new methods for its enhancement. - M. Nauka, 108 p.
CLAIM
1. A method of consolidating the heterosis of a hybrid in subsequent generations, including the use of backcrossings with absolutely homozygous males, characterized in that in order to preserve favorable genes that determine heterosis in the genotype of the hybrid, and at the same time to remove lethals and semi-lethals, backcrossings of hybrids with those obtained from them are used monospermic androgenesis by androgenetic absolutely homozygous males and then, after several backcrosses, switch backcross generations to conventional mass bisexual reproduction through intrahybrid crosses. 2. The method according to claim 1, characterized in that absolutely homozygous male silkworms are obtained by the method of monospermic androgenesis, performed by irradiating eggs in the female’s body with rays at a dose of 80 cr, subsequent mating with males of the original individual hybrids and heating the irradiated inseminated eggs in aged 60 - 80 minutes in water heated to 38 o C for 210 minutes. 3. The method according to claim 1, characterized in that, in order to sharply increase the heterosis of hybrids for industrial use, heterosis is fixed only in individual hybrids arising from two parents that have shown maximum heterosis in comparison with other simultaneously tested hybrids. 4. The method according to claim 1, characterized in that the method of fixing heterosis is used on hybrids of agricultural plants in which it is possible to obtain androgenetic absolutely homozygous individuals by varying methods known for each species of stimulating pollen to embryonic development and converting the germ cells developing from it into absolutely diploid homozygous cells that develop into fertile plants.The method is intended for use in agriculture. The goal is to significantly increase the efficiency of crop hybridization. Heterosis of hybrids appears only in the first generation. It has been established that the attenuation of heterosis in subsequent generations of the hybrid is mainly due to the transition of recessive lethals, semilethals and subvitals into a homozygous state and disruption of a complex of favorable coordinately acting genes. Elimination of these phenomena leads to the consolidation of heterosis in subsequent generations. It is carried out by backcrossing a hybrid with artificially obtained absolutely homozygous androgenetic sons, after which the genetically transformed hybrid is almost completely cleared of harmful genes and at the same time preserves intact the complex of favorable genes that determine heterosis. This makes it possible to completely preserve heterosis in subsequent industrial generations obtained as a result of simple intrahybrid crossings, which has been proven by experiments on silkworms. The method is also intended for agricultural plants, in which it is possible to obtain androgenetic absolutely homozygous individuals. 3 salary, 4 ill.
The invention relates to methods used in agriculture. A well-known natural way of preserving heterosis in subsequent generations is through vegetative propagation in plants that additionally have sexual reproduction. Numerous studies in this area on other plants that are not capable of vegetative propagation and animals were not completely successful (1), because the nature of heterosis was still a big genetic mystery (2). The literature has not even expressed any real theoretical approaches to a radical solution to this important problem. In some animals, heterosis can be fixed by cloning. However, only a few offspring identical to the mother are obtained using this method. In the silkworm, cloning has been developed more successfully, but for practical use in terms of preserving heterosis, it is not acceptable for two reasons: due to the high labor intensity of mass production of parthenogenetic offspring and the lower productivity of the female sex, which makes up the clones, compared to males (3 ). Promising results were obtained by the authors after they developed the method of meiotic parthenogenesis and obtained absolutely homozygous male silkworms from parthenogenetic clones (4). Their backcrossing with a parthenogenetic clone of hybrid origin made it possible to consolidate heterosis in backcross generations (5). But this was the discovery of only the fundamental possibility of fixing heterosis. This method had no practical significance and, therefore, could not be patented as a method. This was explained by the fact that homozygous males could only be obtained from highly viable female parthenoclones with a high propensity for parthenogenesis. It was practically impossible to obtain absolute homozygotes in commercial breeds and hybrids, so meiotic parthenogenesis was used only for reconnaissance experiments aimed at determining the possibility of solving the problem. The invention of a method for fixing heterosis in the silkworm, suitable for production, became possible after the authors discovered monospermic androgenesis (1998, unpublished). The essence of the invention. Heterosis appears only in the first generation of the hybrid. In subsequent generations, starting from the second, it sharply fades. Therefore, in order to grow a heterotic hybrid, it is necessary to repeat intervarietal or interbreed hybridization each time. This process is technically complex and very labor-intensive, and for many plant crops it is simply not feasible, although their hybrids, if they were obtained, would give surprisingly high yields compared to the parent forms. This is exemplified by many agricultural plants. These problems would be radically solved if it were possible to develop an effective way to consolidate heterosis in subsequent generations. This method would simultaneously open up a completely new approach to creating hybrids that are even more outstanding in heterosis. It is known that any industrial hybrid is obtained by crossing a huge mass of individuals of two parental forms. And these individuals are very differentiated in their combinative ability. Therefore, production is content with average heterosis for all individual hybrids taken together, each of which comes from the germ cells of two parents. While rare individual hybrids have truly fantastic heterosis, it is irretrievably lost in the next generation. The proposed method will make it possible to consolidate this powerful heterosis in subsequent generations of the hybrid and propagate it in unlimited quantities. One of the reasons for heterosis was considered to be the beneficial effect on the development and vital activity of the organism of heterozygosity of all genes in general, regardless of their specificity (the “overdominance” hypothesis). Using silkworms, the authors experimentally proved that heterosis occurs as a result of two main reasons. The first is the integration in the genotype of hybrids of a large number of favorable genes, coordinated in their action, that control viability. The second is the transition to a heterozygous state not of all genes of the genotype, but only of recessive details, hemilethals and subvitals (4). In fig. 1 provides evidence of this. Consequently, the decrease in heterosis in subsequent generations of hybrids is mainly explained by the inevitable transition of some recessive parts and semi-lethals into a homozygous state when crossing a hybrid within its limits and the disruption of a complex of favorable genes that increase viability during meiosis. Therefore, the authors came to the conclusion that it is possible to consolidate heterosis in subsequent generations if the complex of all favorable genes in the hybrid genotype is completely preserved or even improved and the recessive lethal and semi-lethal ones are almost completely removed from the genotype. This problem was solved by the authors in the following way. Two genetically distant breeds are selected as the starting material, from crossing which the most highly heterotic hybrids arise. These two breeds produce a series of individual hybrids, each of which comes from only two parents. Through comparative tests, the 10 best individual hybrids in terms of heterosis are selected. From each hybrid, absolutely homozygous descendants are obtained by the method of monospermic androgenesis, which is available to breeders. For this purpose, uninseminated females of any breed are irradiated with rays at a dose of 80 kr. The females then mate with males of the individual hybrids. Layed eggs at the age of 60-80 minutes after laying at a temperature of 25 o C are heated for 210 minutes in water heated to 38 o C. The overwhelming majority of absolute homozygotes die at different stages of development due to the fact that in the haploid genotype they inherited from father, contains many lethal, semi-lethal and subvital genes. When the pronucleus nucleus diploidizes, they pass into a homozygous state, which is most often incompatible with the normal development of the organism. Only those homozygotes survive who, during meiosis, did not receive or received, but very few, harmful genes, mostly of weak effect (5). The grown absolutely homozygous individuals are backcrossed with the original hybrid, thus obtaining the first backcross generation (Fig. 2). The maturation of the original hybrid and absolute homozygotes must be synchronized by delaying the start of growing the first for a time equal to the duration of the development cycle of the selected object. Simple calculations show that in backcross offspring new homozygotes of genes with strong harmfulness cannot appear, and homozygotes of subvital genes, if they were not eliminated from the surviving homozygous androgens, are suppressed by a complex of favorable genes inherited from the original hybrid. This is why heterosis persists in all backcross generations (Fig. 3). The first and subsequent backcross generations are dealt with in exactly the same way as with the original hybrid (Fig. 2). Further backcrosses lead, firstly, to the almost complete removal of details and semi-lethals from the hybrid genotype and, secondly, to the preservation of the numerically predominant part of the genes that provided heterosis in the original hybrid. After 5 or 6 backcrosses, the hybrid, cleared of harmful genes, is massively propagated by intrahybrid crossing. In the offspring obtained as a result of such reproduction, heterosis not only remains at the level of the original hybrid, but even increases somewhat (Fig. 4), which indicates a complete solution to the problem of fixing heterosis in the silkworm. The complete commonality of the genetic basis of heterosis and its attenuation in animals and plants allows this invention to be recommended for consolidating heterosis in agricultural plants, in which it is possible to obtain absolutely homozygous individuals of androgenic origin from hybrids. They are obtained by stimulating the embryonic development of haploid pollen, followed by the transformation of its germ cells into diploid ones, which develop into viable fertile plants. The technique varies depending on the biological characteristics of the crop. Graphic materials. Fig. 1 A. A direct relationship is shown between the yield of silkworm cocoons - the main indicator of heterosis (1) and the levels of heterozygosity (2) of genetic variants of the hybrid that are not purified from lethals and semi-lethals. The yield and heterozygosity indicators of the original hybrid of the first option (1) are taken as 100%. B. It has been shown that there is a complete absence of dependence between the cocoon yield (1) and the levels of heterozygosity (2) in genetic variants purified from lethals and semi-lethals. This proves the inconsistency of the “overdominance” hypothesis of heterosis and the possibility of maintaining heterosis in backcross generations. Fig. 2. Scheme for purifying silkworm hybrids from recessive lethals and semi-lethals through backcrossing of hybrids with absolutely homozygous males of breed A and B obtained from them. F 1, F 2 - hybrid of the first and second generation. F b1, F b2 - first and second backcross generations. Fig. 3. Viability of the original hybrid (1) and backcross generations (II), obtained according to the scheme presented in Fig. 2. Fig. 4. Demonstrates indicators of the frequency of harmful genes in the heterozygous state (1), cocoon mass (2), viability (3) in the original hybrid (I) and the transformed hybrid after four consecutive backcrosses with homozygous males (II), as well as in three consecutive inbred generations (III-V). Each genetic variant was reared simultaneously with a control parthenogenetic hybrid, the indicators of which were taken as 100%. In all genetic variants, heterosis is higher than in the original hybrid, which indicates a radical solution to the problem of fixing heterosis. The stable preservation of heterosis in all backcross generations has already indicated the fundamental effectiveness of the developed method. But backcross generations are not practical due to the difficulty of obtaining them. Therefore, in the final experiment on silkworms, they studied the possibility of fixing heterosis not in backcross, but in normal generations. In this final experiment, the original hybrid was first subjected to four backcrosses with homozygous males. As a result, the frequency of heterozygotes for lethals and semi-lethals decreased to 6.2% from 100% in the original material. Next, backcross generations were propagated by inbreeding. Each inbred generation was obtained by crossing a brother with a sister within each individual family. As a result, the frequency of harmful genes suppressed by normal alleles decreased in the first inbred generation to 4.7, and in the second and third - to 3.5 and 2.6%, respectively. Inbred reproduction has an extremely detrimental effect on all economic indicators of normal inbred offspring. But in our experiment, it not only did not have a depressing effect on the inbred offspring, but, on the contrary, led to an increase in its average weight of one cocoon and viability compared to the original, control hybrid (Fig. 4). Consequently, the problem of fixing heterosis in hybrids of subsequent generations has been radically solved. BIBLIOGRAPHICAL DATA 1. Inge-Vechtomov S.I. 1989. Genetics with the basics of selection. M. "Higher School", on page 557. 2. Hutt F. 1969. Animal genetics. Per. from English edited by Doctor of Biology Sciences Ya.L. Glembotsky. M., "Spike", on page 322. 3. Strunnikov V. A. 1998. Cloning of animals: theory and practice. - Nature, N 7, pp. 3 -9. 4. Strunnikov V.A. 1987. Genetic methods for selection and sex regulation of silkworms. M. VO "Agropromizdat", on page 35. 5. Strunnikov V.A. 1994. The nature of heterosis and new methods for its enhancement. - M. Nauka, 108 p.
Claim
1. A method of consolidating the heterosis of a hybrid in subsequent generations, including the use of backcrossings with absolutely homozygous males, characterized in that in order to preserve favorable genes that determine heterosis in the genotype of the hybrid, and at the same time to remove lethals and semi-lethals, backcrossings of hybrids with those obtained from them are used monospermic androgenesis by androgenetic absolutely homozygous males and then, after several backcrosses, switch backcross generations to conventional mass bisexual reproduction through intrahybrid crosses. 2. The method according to claim 1, characterized in that absolutely homozygous male silkworms are obtained by the method of monospermic androgenesis, performed by irradiating eggs in the female’s body with rays at a dose of 80 cr, subsequent mating with males of the original individual hybrids and heating the irradiated inseminated eggs in aged 60 - 80 minutes in water heated to 38 o C for 210 minutes. 3. The method according to claim 1, characterized in that, in order to sharply increase the heterosis of hybrids for industrial use, heterosis is fixed only in individual hybrids arising from two parents that have shown maximum heterosis in comparison with other simultaneously tested hybrids. 4. The method according to claim 1, characterized in that the method of fixing heterosis is used on hybrids of agricultural plants in which it is possible to obtain androgenetic absolutely homozygous individuals by varying methods known for each species of stimulating pollen to embryonic development and converting the germ cells developing from it into absolutely diploid homozygous cells that develop into fertile plants.
Detailed solution to paragraph § 32 in biology for 10th grade students, authors V.I. Sivoglazov, I.B. Agafonova, E.T. Zakharova. 2014
Remember!
What is selection?
Give examples of animal breeds and plant varieties known to you.
Antonovka apple varieties, Severyanka pear, dog breeds: Rottweiler, miniature poodle, collie.
Review questions and assignments
1. What is selection?
Selection (from Latin selectio - selection) is the science of creating new and improving existing plant varieties, animal breeds and strains of microorganisms. At the same time, selection is understood as the process of creating varieties, breeds and strains. The theoretical basis of selection is genetics.
2. What is called a breed, variety, strain?
A breed, variety or strain is a collection of individuals of the same species, artificially created by man and characterized by certain hereditary properties.

5. What difficulties arise when performing interspecific crosses?
Distant hybridization involves crossing different species. In plant growing, with the help of distant hybridization, a new grain crop has been created - triticale, a hybrid of rye and wheat. This crop combines many of the properties of wheat (high baking qualities) and rye (the ability to grow on poor sandy soils). A classic example of interspecific hybrids in livestock breeding is a mule, obtained by crossing a donkey with a mare, which is significantly superior to its parents in endurance and performance. In Kazakhstan, by crossing wild mountain argali sheep with fine-wool sheep, the famous argali sheep breed was created. However, the use of interspecific crosses has certain difficulties, because the resulting hybrids often turn out to be infertile (sterile) or low-fertility. The sterility of hybrids is associated with the absence of paired homologous chromosomes. This makes the conjugation process impossible. Therefore, meiosis cannot be completed and no germ cells are formed.
6. Are interspecific hybrids produced and used in your region? Using additional sources of information, find out which species are hybrids of organisms such as bester, honorik, hinny, and raphanobrassica. What interest are they for agriculture?

Think! Remember!

2. Why do each region need its own plant varieties and animal breeds? What varieties and breeds are typical for your region? What are their features and advantages?
Since environmental conditions are different in different regions, varieties and breeds must be adapted to specific conditions. Features of crop production in the Southern Urals

3. Of the wide variety of animal species living on Earth, humans have selected relatively few species for domestication. What do you think explains this?
The process of domesticating wild animals begins with the artificial selection of individual individuals to produce offspring with certain characteristics necessary for humans. Individuals are typically selected for certain desirable characteristics, including reduced aggression towards humans and members of their own species. In this regard, it is customary to talk about taming a wild species. The purpose of domestication is to use an animal in agriculture as a farm animal or as a pet. If this goal is achieved, we can talk about a domesticated animal. The domestication of an animal radically changes the conditions for the further development of the species. Natural evolutionary development is replaced by artificial selection based on breeding criteria. Thus, as part of domestication, the genetic properties of the species change.
4. Heterosis usually does not persist in subsequent generations and fades away. Why is this happening?
When crossing different breeds of animals or plant varieties, as well as during interspecific crossings in the first generation, the viability of hybrids increases and powerful development is observed. The phenomenon of superiority of hybrids in their properties of parental forms is called heterosis, or hybrid vigor. It appears in the first generation, and fades away in the second.
5. Why do you think ligers are born only in zoos and are not found in the wild? Explain your point of view.
Ligers - interspecific hybrids between a lion and a tigress - look like huge lions with blurred stripes. Therefore, his parents belong to the same biological genus of panthers, but different species. In appearance, it is noticeably different from its opposite hybrid, the tigrol. It is the largest representative of the cat family currently existing. Looks like a giant lion with blurred stripes. Ligers are not found in the wild mainly because lions and tigers have little chance of meeting in the wild: the lion's modern range includes mainly central and southern Africa (although India has the last surviving population of Asiatic lions), while the tiger exclusively Asian look. Therefore, crossing of species occurs when animals live for a long time in the same enclosure or cage (for example, in a zoo or circus), but only 1-2% of pairs produce offspring, which is why there are no more than two dozen ligers in the world today.
6. Do you think mass selection can be used when breeding animals? Prove your opinion.
Not used. mass selection is selection by phenotype. Individual - by genotype. Producers in animals are individuals with a well-defined pedigree, i.e. the genotype for the desired traits is quite well known. And the characteristics of animals - it takes time to reach sexual maturity, a small number of offspring (compared to plants - now can be considered a solved problem - artificial insemination, surrogate females) and the impossibility of asexual reproduction.
7. Using additional literature and Internet resources, prepare a message or presentation about the history of selection from ancient times to the present.
Selection as a method of developing breeds of domestic animals and varieties of cultivated plants has existed for a long time. About 8000-9000 years ago, with the advent of agriculture in the Middle East, and later in Europe and Asia, the development of crop and livestock farming began. Since that time, people began to engage in artificial selection in order to breed animal breeds and plant varieties with economically valuable qualities. The first breeding activities, known almost 6000 years ago in Elam (Mesopotamia), can be judged by the image of the pedigree of horses found on a signet. There is also information that the Arabs, long before the new era, used artificial pollination of date palms. The Roman Empire preserved documents detailing the techniques used in breeding animals. In the works of scientists of Ancient China and Ancient Rome, there are indications of the importance of selecting ears in cereals and recommendations are given for carrying out such selection.
At first, breeding activities were limited to selection. It was unconscious in nature and lasted for a long time (10-15 years). Breeders, without a theoretical basis, were guided by experience and intuition. They took into account the beneficial properties of the parental individuals, but could not carry out targeted selection. The results of crossing often turned out to be unexpected, and the expected trait was not found in the offspring. Nevertheless, unknown breeders left a legacy of many valuable varieties of cultivated plants and breeds of domestic animals. For example, a number of the best varieties of cotton now cultivated in Russia and the USA were borrowed from the peasants of old Mexican villages. Using the method of unconscious selection, varieties of fiber flax were bred in some areas of Pskov: low-growing plants were used for household needs, and seeds of tall ones were used for sowing. There are known varieties of winter (for example, Krymka, Poltavka, Sandomirka) and spring (Ulka, Girka, Syr-Bidai, etc.) wheat with valuable economic qualities, bred in ancient times.
However, selection for economically useful traits and properties without taking into account the mechanisms of their heritability and variability often gave undesirable results. For example, selection for the appearance of fine-wool sheep for polling led to the appearance of cryptorchidism; getting rid of piebald hair on the neck of Romanov sheep weakened their vitality; An increase in hair growth in sheep was accompanied by a decrease in their weight. It was not possible to develop a pure line of Wyandottes (a breed of chickens) with a rose-shaped comb; Despite the culling of chicks with leaf-shaped combs, they appeared in the offspring. Obviously, the breed consisted of generalozygotes for this gene, since homozygotes had reduced fertility.
All this indicated that the desired result cannot be obtained without theoretical knowledge. From the end of the 18th - beginning of the 19th century. The work of breeders was already of a scientific nature. The main task of breeding was the study of the genetics of such traits as animal productivity and plant yield. Solving selection problems is impossible without knowledge related to genetic analysis, i.e. without knowledge of the type of inheritance of traits (dominant or recessive), the type of dominance, the nature of inheritance (autosomal or sex-linked, independent or linked), the type and nature of gene interaction in ontogenesis . Breeders should pay main attention to the problems of the relationship between the genotype and the environment, since the expressivity and penetrance of the studied traits largely depends on the factors of the latter.
8. Are there breeding stations or centers in your region? What research are they doing? What are their achievements? Together with your teacher, organize an excursion to such a station.
Yu-U Research Institute of Horticulture and Potato Growing, Chelyabinsk
Eh, apple, ranetochka...
In 1931, on the initiative of I.V. Michurin, the first scientific research institution for horticulture in the Southern Urals was created - the Ural Zonal Fruit and Berry Experimental Station. The organizer of this station was Valery Pavlovich Yarushin.
And the very next year, scientific research began on the selection and selection of varieties suitable for cultivation in the harsh conditions of the Chelyabinsk and Kurgan regions, including the then Kamyshlovsky and Kamensk-Uralsky regions of the now Sverdlovsk region. Scientists began to survey and collect the best forms of fruit and berry crops east of the Ural Range. In 1934, station employees registered a large tract of wild cherries - 2270 hectares - in the Karagai forest dacha (Annensky Bor). In the same year, a scientific expedition led by Doctor of Agricultural Sciences E. P. Syubarova (BelSRRI of Fruit Growing) and Chelyabinsk scientist M. N. Salamatov examined thickets of wild steppe cherries in the Verkhneufaleysky and Poltava districts of the Chelyabinsk region. At the same time, extensive collections of plums were imported from the Far East, Canada, North America, central Russia, the Volga region, and Karzin plums from Siberia. From the selected material in 1937, scientists identified varieties and proposed the first Ural assortment of berry crops.
In those years, both among the people and agronomic science, it was believed that due to the harsh climate with frosty and long winters, gardening in the Urals was impossible. It took South Ural scientists only twenty years - by breeding standards, a very short period of time - to refute this widespread opinion. Thanks to the selection and study of new varieties of fruit and berry crops at the Chelyabinsk experimental station, horticulture began to develop rapidly in our country.
The first collective gardens in the region appeared soon after the war. In 1948, Traktorosad, Druzhba in the Metallurgical District, Lokomotiv in the Sovetsky District, and the gardens of the Magnitogorsk Iron and Steel Works were formed. Their appearance was preceded by a long and viscous struggle in the corridors of the then government with opponents of the creation of such gardens. Nevertheless, collective gardening has developed and is still developing. Currently, gardeners produce the bulk of fruits and vegetables.
By the beginning of the 50s, our scientists collected and studied 442 apple tree varieties, including 210 varieties of Ural-Siberian selection. The first varieties of fruit and berry experimental station officially issued with copyright certificates were apple trees. The basis of the pre-war assortment was local ranetki. They are distinguished by their adaptability to the Ural and Siberian climate, high yield and small fruit size weighing from 15 to 50 g. The older generation still remembers these bright, beautiful Ranetka apples - Lyubimets, Anisik Omsky, Ponikloe. They seemed like the ultimate dream then, especially for children. But by the 60s, the institute already had 25 zoned varieties: 14 apple trees, 4 pears, 4 plums and 3 berry crops.
In 1964, the Ural Zonal Fruit and Berry Experimental Station was renamed the Chelyabinsk Fruit and Vegetable Breeding Station named after. I. V. Michurina. By that time, the specialized trust “Plodoprom”, headed by Vsevolod Ivanovich Nazarov, was already successfully operating in the region. Close cooperation between science and production, the passion and enthusiasm of their employees have raised South Ural horticulture to unprecedented heights. With the support of regional leaders, fruit nurseries were created: "Smolinsky" in the suburbs of Chelyabinsk, "Michurinsky" in Kartaly, "Raduzhny" in Magnitogorsk, "Tyubelyassky" in the mining zone. State variety testing sites were organized at the Smolinsky and Michurinsky fruit nurseries.
It was then, in the 50-60s, that small gardens from 20 to 100 hectares were established on collective and state farms located under the MTS. Some of them have survived to this day. At the peak of the development of horticulture in the region, public gardens occupied an area of 8 thousand hectares, and the industry itself was generally profitable.
In the fifties, work on scientific support for potato growing also began in our region. By the beginning of the 80s, the Chelyabinsk fruit and vegetable breeding station named after. I.V. Michurina had 29 of her own varieties of fruit and berry crops and potatoes, included in the State Register of Breeding Achievements approved for use.
Station scientists began to use the world collection of forms of fruit and berry crops and potatoes from the All-Union Institute of Plant Growing and other research institutions, including foreign ones, in plant breeding. The genetic fund was involved in the selection process, new breeding technologies were introduced that made it possible to shorten the selection process, and the training of highly qualified personnel increased. The scientific developments of South Ural scientists began to be implemented in the farms of the Chelyabinsk, Kurgan, Kustanai, Orenburg and other regions, in the Republic of Bashkortostan.
New name - new goals
In November 1991, by decision of the Government of the Russian Federation, the Chelyabinsk Fruit and Vegetable Experimental Station named after. I.V. Michurina was transformed into the South Ural Research Institute of Fruit and Vegetable and Potato Growing. Not only the name has changed. More serious goals and directions of research activities were set for the institute. In addition to the selection of fruit and berry crops and potatoes, life required the creation of new resource-saving, environmentally friendly technologies for the selection and cultivation of these crops, scientific research on hybridization, as well as the production of elite seedlings of new promising varieties of horticultural crops and potato seeds on a healthy, virus-free basis using biotechnology.
Over the entire period of the institute’s activities, starting with the Ural Zonal Fruit and Berry Station, Chelyabinsk breeders have created more than 200 varieties of fruit and berry crops, 18 varieties of potatoes, developed technologies for industrial and amateur gardening, technologies for the production of potatoes with a yield of up to 70 t/ha.
Over the years, 110 varieties were included in the state register of selection achievements approved for use. The level of research work is evidenced by the fact that today the institute has more than 100 copyright certificates and patents for varieties and inventions.
For the first time in the Southern Urals, models of intensive varieties of fruit and berry crops, potatoes until 2020 have been created, selection schemes have been developed for productivity, winter hardiness, incl. resistance of flowers to spring frosts, product quality, immunity. The Institute has a rich genetic fund of horticultural crops, which includes 64 thousand hybrid plants, incl. 39 - fruit crops, 25 thousand seedlings of berry crops. The volume of hybrid crosses amounts to 45 thousand flowers.
Scientific research on selection and agricultural technology of horticultural crops and potatoes is carried out in creative collaboration with leading research institutes and experimental stations in Russia, near and far abroad.
Today the institute has a breeding garden with an area of more than 100 hectares. It is here that the main scientific activity of South Ural breeders is concentrated.
Apple tree. For the first time in the world practice of horticulture, the institute's breeders, using an originally developed method, developed varieties of natural dwarfs with a tree height of 1.5-2.5 m. When propagated on clonal vegetatively propagated dwarf rootstocks, they become natural dwarfs (0.8-1.5 m).
Pear is an amazing crop in the Urals. Fruits every year. The fruits are tasty, sweet, suitable for processing. Involvement of selected forms of Ussuri pear in breeding made it possible to create varieties characterized by high winter hardiness and productivity, high taste qualities of fruits, monogenic resistance to scab and field resistance to pear gall mite.
Apricot and plum. Local varieties have been bred and selected forms have been identified, characterized by increased winter hardiness of fruit buds and high quality fruits.
Cherry. Experimental data have been accumulated to create genotypes resistant to coccomycosis. Isolation of donors with a gene for mono-resistance to coccomycosis and identification of forms of steppe cherry with field resistance to coccomycosis continues. For this purpose and to replenish the collection, expeditions were carried out to examine wild cherry plants in the territory of Bashkiria, Chelyabinsk and Kurgan regions. Forms of steppe and forest cherries were selected that were resistant to coccomycosis, large-fruited, and with good fruit taste.
Berry crops. Research is being conducted on the selection of new varieties that are resistant to adverse environmental factors, highly winter-hardy, with increased resistance of flowers to spring frosts, highly productive, and with excellent fruit quality. Models of the optimal variety of berry crops have been compiled, taking into account the technological requirements of breeding for 2020-2025. Technologies for cultivating rose hips, gooseberries, honeysuckle, currants, and technologies for propagating currants in film greenhouses have been developed.
Bring back its former glory
The reforms carried out in the Russian agricultural sector since the early 90s have turned out to be destructive for horticulture. As a branch of the national economy, it virtually ceased to exist. Only in the Chelyabinsk region the share of industrial gardening decreased from 65% to 1%. There are almost no fruit-bearing plantings left, experimental production fruit nurseries have practically ceased their activities, and their products satisfy market demand by only 12-15%.
There is no targeted program for the development of industrial horticulture in the region, based on reliable materials from the inventory of plantings and new developments of scientific institutions in recent years.
This is not science's fault. Scientists from the South Ural Research Institute of Horticulture and Potato Growing have more than once proposed to form the scientific basis for the revival of the industry. However, the state of the macroeconomy and bureaucratic conservatism do not provide an opportunity to move forward. The expectation that collective amateur gardening (i.e., the private sector) will make up for the losses of industrial gardening is not justified. Yes, and he cannot justify himself. Despite the fact that UUNIIPOK annually produces more than 60 thousand seedlings of fruit and berry crops for the population, the private sector suffers from a lack of planting material, especially new varieties, the need for which is met by 47-50%. Small producers of seedlings - private traders - are trying to fill this niche. But the quality of their planting material often does not stand up to criticism in all respects.
Is it realistic to revive community gardening in the new economic conditions?
Quite, say South Ural scientists and breeders. But for this, at the state and regional level, in their opinion, it is necessary to resolve the most pressing issues:
From the areas under gardens from the beginning of territory preparation until the plantings begin to bear fruit, cancel the land tax
Allocate capital investments for planting perennial plantings on a repayable basis
At industrial enterprises in the region, organize the production of special equipment for gardening
Laws are needed to protect domestic producers in their own market
Encourage small businesses to organize processing of horticultural products
In the vocational education system it is necessary to organize training for horticulture
The unification of the scientific potential of Ural agricultural scientists is also very important. For these purposes, it is necessary to create a Ural Scientific and Methodological Center with its location in Chelyabinsk. The benefits of such a union are enormous. Proof of this is the potato coordinating council, created in April 2000 at the initiative of the State Scientific Institution YuUNIPOK on a voluntary basis. Over the 8 years of joint work, potato growing institutes have carried out a powerful mobilization of the gene pool, expanded collections, agreed on crossing combinations, and regularly exchange information, source and breeding material. The practice of the coordinating council has shown that this form of work is extremely effective, viable and therefore deserves development and improvement.
Work has also begun to unite horticultural institutions. A multilateral agreement was signed between six scientific institutions of the Ural and adjacent regions - State Scientific Institution YuUNIIPOK, BashNIISKH, Udmurt Research Institute of Agriculture, Kostanay Research Institute of Agriculture, Kazakh Research Institute of Agriculture, Karabalyk Experimental Station.
Agricultural science today is going through difficult times, but it is alive and ready to contribute to the revival of agricultural production, including the South Ural industrial horticulture.



